 您当前的位置:
您当前的位置:
研究老鼠舔舔舔,意义是什么呢?
2022年3月9日,约翰霍普金斯大学Daniel H。 O’Connor团队(第一作者为许多博士)在Nature上发表了文章Cortical processing of flexible and context-dependent sensorimotor sequences。

“你研究老鼠舔舔舔,意义是什么呢?”我的朋友问。
每次给报告讲这次的工作,我都会放一段网红小姐姐用舌头打音游的短视频。贝八钢琴奏鸣曲响起,音符从手机屏幕上方快速落下,只见美颜滤镜拉满的小姐姐伸出舌头,在手机屏幕上快速且精准地左右点触,音符在准确的时间、准确的位置被完美击破。然后我会说:“大家看,我训练小鼠做的事情,人类也干,所以四舍五入我相当于研究人了!”当然,这只是个活跃气氛的玩笑。
但不难发现,很多人经常忽视我们的舌头其实是人最灵巧的运动器官。灵巧到什么程度?我们随随便便说一句话就会包含十几、几十个元音和辅音,发每一个音都需要舌头和其它发声结构摆出一个不同的“姿势”(gesture),一系列姿势要在短短几秒钟内飞快地完成(每秒约7个语音(phones))。如果把舌头类比拉小提琴的左右手,屏幕前的读者朋友们可以说是人均“帕格尼尼”,语速稍快就“《野蜂飞舞》”了(每秒约10个音符)。舌头不仅极致灵巧,控制舌头的脑环路还有极强的学习能力或可塑性。陌生的一系列动作通过练习会变得熟练,直至了熟于胸。最近互联网上有位因工伤瘫痪的小哥,凭着不懈的努力学会了用嘴操作控制器打游戏,这也许是舌头运动序列可塑性这个严肃科学问题最让人暖心的例子了吧。
一切行为最终都需要通过神经细胞的活动来控制实现。不同行为序列的具体表现形式虽然不同,它们背后也许共享着神经环路层面的基本原理。深入研究这些原理需要使用恰当的模式生物,以及和问题相匹配的行为范式。小鼠凭借着丰富的电生理、影像学、遗传学、解剖学工具库和数据库、以及相对低廉的操作成本,已经成为了研究认知与行为背后神经环路原理的重要手段。然而在此工作之前,小鼠并没有一个能够充分体现复杂运动序列特征的行为范式,使得相关研究难以深入。
老鼠虽然不会说话,但它们非常擅长使用牙口舌鼻胡须等面部器官。就像小姐姐用舌头玩音游,我们在实验中训练小鼠舔面前的一系列位置,有的从左向右,有的从右向左,有循回往复。和音游不同的是,小鼠的面前没有一个显示“音符下落”的手机屏幕或者任何其它外界提示,它们能感知的只是自己舔到了还是没舔到的触觉反馈。经过训练,业务熟练的小鼠可以每秒舔过约7个给定位置,和人说话时舌头“姿势”的变化速度可谓旗鼓相当。
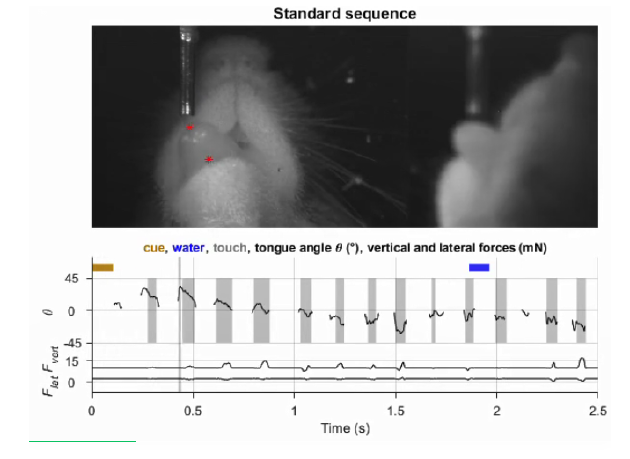
因为这是个全新的行为范式,我们一上来并不确定哪些脑区会参与控制口部的序列运动,于是决定一边让小鼠舔舔舔,一边用光遗传的手段对额叶和顶叶进行一系列局部的定点抑制。抑制额叶的前侧运动皮层(包括ALM和舌颚M1)会导致小鼠无法启动序列,或者显著减弱舌头的运动能力;抑制初级感觉皮层的舌、颚代表区(舌颚S1)会打乱序列,但不会减弱运动能力;最让人意想不到的是,如果小鼠做完了一系列舔正要结束,此时抑制舌颚S1或初级运动皮层的身体代表区(体M1)反而会让小鼠停不下来。(如果有条件,建议观看原文附带的实验视频以获得更直观的体会。)
锁定了舌颚S1、舌颚M1、和ALM这三个关键区域之后,我们用硅基电极阵列进行了单细胞分辨率的电生理记录。神经环路处理信息往往以神经元集群为单位,所以接下来我们仔细讲讲在集群层面不同脑区对运动序列信息的编码和处理。
复杂的运动序列有一个重要特征——层级(hierarchy)。比如游戏中的每一个具体操作都对应着一个特定的键位,一套连招需要多个键位顺序组合,而一次战斗又需要多个键位和连招在更大的时间尺度上进行排列。不仅如此,当对战态势出现变数,玩家还需要灵活调整。人类的语言也有明显的层级结构,比如音节组成词语,词语构成句子,句子需要根据说话者的意图灵活组合。
解码分析发现,从舌颚S1到舌颚M1再到ALM,神经集群所承载的信息层级不断提升。舌颚S1和舌颚M1神经集群更多处理舌头实时的“姿势”,包括长度、速度、侧偏角度;而ALM则更多承载了隐含(latent)的信息,包括目标方位、正在完成哪个序列、离序列终点的奖励还有多远。这些抽象的隐含信息不表现在具体动作上,它们是驻留在脑子里起组织作用的信号。
比如为什么要知道自己在执行哪一个序列呢?请读者设想自己从1数到10,或者从10数到1。每当我们念出5,接下来是4还是6需要我们心里知道自己是在正数还是在倒数。实际上对于任何存在层级的行为序列,不管是游戏、舞蹈、还是语言等等,序列类型的信息都决定了子序列能否正确组合。但是小鼠的脑子会有这种能力吗?为了让实验足够有说服力,我们为小鼠设计了比1到10、10到1还要复杂的“Boss”级任务。如果把不同的目标位置编上号,那么小鼠需要不断地交替完成1-2-3-4-5-4-3-2-1-2-3-4-5和5-4-3-2-1-2-3-4-5-4-3-2-1这两个序列。这两个序列里不仅有数字相同但走向不同的情况,还有数字相同、走向也相同、但先后不同的小片段,比如“2-3-4”在第一个序列里位于开头和结尾,第二个序列里则位于中间。实验结果发现,ALM的神经元集群的确在动作完全相同的片段中也会保存序列类型信息。序列类型信号的存在说明小鼠能够跨越三个序列层级进行整合,即单个动作->子序列->总序列。这个发现大大拓宽了未来人们能在小鼠身上研究的可能,进一步回答层级整合的环路与计算机制。
当外界条件发生变化时,大脑需要一枚信号弹,告诉自己原先的序列该重新调整了。在另一组实验中,我们让目标在中途突然跳离预期轨迹,这时候小鼠需要察觉变化,见招拆招。目标有两种跳离方式,它们的方向相反,小鼠做出的应对动作也完全不同。有意思的是,我们观测到舌颚M1和ALM的神经元集群不仅处理具体的运动信息,同时还产生了“信号弹”似的隐含信息。这个信号不编码具体动作,而是抽象地表达了中止某个当前动作序列进而转向某个新序列的需求。它对大脑如何监测偏差(error monitoring)、归纳错误、并调动序列分支的原理有很大启发。
我还是本科生的时候参加过一个国际学术会议的联谊活动,当时啥都不懂的我问几个教授怎么看在小鼠身上研究视觉。“小鼠基本上就是瞎子吧”、“哈哈哈哈哈啊哈”、“还不如用青蛙呢”教授们一边晃着酒杯一边调侃着。然而短短几年之后,随着一系列工具鼠和实验模式的建立与推广,小鼠视觉研究如雨后春笋般俨然霸占了神经科学年会的十几面海报墙。小鼠还是当年那个“瞎子”,但当合适的条件出现,它们的视觉系统仍然可以揭示神经环路的很多基本遗传特征和计算原理。
对于人类来说,因为语言的存在,舌头成为了和内心世界联系最紧密的器官之一。小鼠是个“哑巴”,也不及灵长类的认知能力,但当合适的条件出现,我相信小鼠仍然会是探索环路机制的排头兵、是跨领域跨机构合作的交汇点、是检测新技术新理论的试验田。希望这次的工作能成为这片田地的一粒种子,开向无限可能的未来。
来源:新浪科技综合
南京中医药大学 版权所有